





Abstract
Natural nests of the European ground squirrel (Spermophilus citellus) are constructed almost exclusively of fresh fescue (Festuca pseudovina: Poaceae). We performed laboratory experiments to understand the functional significance of preference of wild squirrels for nest material. We examined the factors contributing to nest quality by monitoring the construction and analyzing the composition of squirrel nests. As in the wild, squirrels showed strong preference for fescue during the laboratory tests and preferred fresh material to dry material. Because preference for fresh material was not expected, because high water content reduces insulation, we manipulated the moisture content of nests made from either fresh or dry fescue to determine how nest quality and moisture level contribute to insulation. We found that the insulation property of fresh grass nests was superior to nests constructed from dry grass only. Intracellular water in the nest material did not affect thermal conductance and insulation significantly. We concluded that fresh fescue provided a more flexible material that allows squirrels to construct nests with better insulation under both field and laboratory conditions.
Nest building has evolved in many taxa but is used mainly by endothermic animals that can maintain body temperature above ambient conditions. Although nests share common features within a taxon, their properties are determined by their function. The insulation quality of nests is dependent on several factors, such as nest structure (McGowan et al. 2004; Redman et al. 1999); height, thickness, and volume (Grubbauer and Hoi 1996; Szentirmai et al. 2005); nest material quality (Mertens 1977); and moisture content (Pinowski et al. 2006).
Ground-dwelling sciurids (Spermophilus spp.) are an abundant group of burrowing mammals, making their long burrow systems and building their nests at various depths in the soil (Hut and Scharff 1998; Michener 2000, 2002, 2004; Mrosovsky 1968; Ruzič 1978). Squirrel burrows and nests serve a number of functions, such as providing shelter during resting, sleeping, or hibernation and protection against unpleasant environmental periods (Hut and Scharff 1998), defense from predators, as a place to store food, and for reproduction (Bethge et al. 2004; Long et al. 2005; Meadows 1991; Pizzimenti and Hoffmann 1973). In general, nests of burrowing small mammals insulate them from fluctuating ambient temperatures (Casey 1981). As a result, the nest can affect the magnitude of energy conservation and consequently is considered crucial (Bethge et al. 2004; Geiser 1988; Houston and McNamara 1993; Lovegrove et al. 2001; McCafferty et al. 2003; Redman et al. 1999) or even critical (Barclay et al. 2001; Pinowski et al. 2006) to survival or reproduction (Lamprecht and Schmolz 2004). Squirrels spend about 7 months underground during the hibernation period (Millesi et al. 1999), when even small differences in insulation can be of functional significance because breeding success is closely linked to body mass at vernal emergence (Millesi et al. 1999).
Nest insulation hardly influences nest temperature during torpor because the very low metabolic rate during torpor results in a minimal temperature gradient between the torpid animal and its surroundings. Therefore energy savings from the nest are usually marginal during torpor itself. However, a good nest will help an individual save energy during arousal and normothermia when energy expenditure is high during the hibernation season. In addition, should a hibernator be forced to thermoregulate during torpor because ambient temperature falls below the optimum (Buck and Barnes 2000; Geiser and Kenagy 1988), the temperature gradient between animal and surroundings will increase, and good nest insulation will reduce energy expenditure.
Nest quality also can contribute to survival in the active season. The relationship of insulation and loss of body mass changes in the active season (i.e., during the mating period), when better insulation quality of the nest can help reduce loss in body mass. Mating or rearing offspring is energetically costly primarily due to decreased foraging time (Millesi et al. 1999); therefore a good nest can save energy, contributing to reproductive success. Our manipulative study on the European ground squirrel (Spermophilus citellus) has shown that to avoid high temperatures (Váczi 2005) ground squirrels hide underground in the humid nest chamber at midday during summer drought periods, possibly to enhance cooling (Váczi et al. 2006). In a field study focusing on plant use for dietary and nest-building purposes (C. I. Gedeon, pers. obs.) we found that squirrel nests contained fescue (Festuca pseudovina) in quantities well above chance level, but contrary to our expectations relative to insulation needs, nests contained fresh green material not the dry hay known to be preferred by similarly sized Mongolian gerbils and pikas (Millar and Zwickel 1972).
The aim of the study was to understand factors contributing to the nest material choice of the European ground squirrel, and to test the consequences of preference for fresh green nest material in nest building as insulation. We asked the following questions: Do European ground squirrels have a specific preference for fescue in nest construction? Does moisture content of the plant affect choice? To what extent does moisture content of the nest material affect nest insulation?
Materials and Methods
Animals and housing.—Our field and laboratory study procedures were part of an approved research permission issued by the Authority of Nature Conservation in charge. In addition, our laboratory procedure also was approved by the Ethical Committee for Animal Experiments at Eötvös University, and followed the rules detailed in guidelines of the American Society of Mammalogists (Gannon et al. 2007). After the experiment, in the spring of 2008, study animals were released into their original habitat.
Eight wild male European ground squirrels were trapped on the grassy airport of Dunakeszi, Duna-Ipoly National Park, Hungary (47°36′51.7″N, 19°8′32.5″E), in September 2007 and were transferred to a climate-controlled room at Eötvös Loránd University, Budapest. Squirrels were housed individually in plastic containers of 15 × 21 × 36-cm size. Containers were open at the top but were covered with a removable wire mesh. Barrel-shaped (16-cm diameter × 8-cm height), opaque plastic boxes were situated in every container as nest boxes with removable tops. The squirrels could enter the nest box through an elbow-shaped tunnel (6 cm in diameter) attached to the side of the box. Room temperature was measured continuously and maintained constant (approximately 25 °C). Illumination (12L:12D, with light on at 0600 h) was constant, and both food (rabbit chow; Galgavit Ltd., Gödöllő, Hungary) and water were given ad libitum during experiments (Németh et al 2009).
Nest material choice.—To determine whether squirrels selectively choose certain materials for nest building, 2-way nest material choice tests were conducted where we tested the species preference (experiment 1) and the moisture content preference (experiment 2) in nest building. For these experiments F. pseudovina (pseudovina fescue) and Bothrio-chloa ischaemum (yellow bluestem) plant tussocks were collected on the grassy airport of Dunakeszi, because these were the dominant species of the grassland and the main constituents of natural squirrel nests (C. I. Gedeon, pers. obs.). Consequently, animals were familiar with the plants provided for nest construction.
Nest-building behavior of the animals was recorded in their home cages by a continuous and computerized digital video system (Cameras PTC-150S; Elmo Ltd., Nagoya, Japan) positioned 2.5 m above the cages. In both experiments 2 grass tussocks of equal volume (2 dm3) and known composition (dried Festuca versus dried Bothriochloa in experiment 1, and fresh, humid Festuca versus dried Festuca in experiment 2) were presented to the animals for nest building in the early morning, and their behavior was recorded continuously. When one-half of the volume of the grass was used the test was terminated. At that point possible difference in grass usage should have been maximal because if only fresh or dry grass had been used for nest building, squirrels would have switched to the other tussock if they had required more grass for nest construction. Consequently, the fresh:dry grass ratio would have been biased due to limited volume of grass resources.
The position of the 2 grass tussocks was assigned randomly to one of the opposite corners of the cages, with a distance of 25–35 cm between the tussocks. From the video recording we quantified the nest-building behavior between 0900 and 1200 h (3 h), which is considered to be the main period for surface activity of these squirrels in the wild (Katona et al. 2002; Váczi et al. 2006). The time duration (min) spent within 10 cm from the 2 grass tussocks was quantified from the video material as a measure of nest material collection.
Subsequently, assembled nests were removed from the nest boxes. From each nest 3 independent samples were picked with forceps, each containing 25–40 pieces of grass, and these samples were analyzed with a microscope using 40 × magnification (Petróczi and Altbäcker 1994). In the analyses we determined the grass species (experiment 1) and, based on the color of the grass leaves, whether they were fresh (humid, dark-green) or dry (light-green) material (experiment 2). The average of the 3 samples was subjected to statistical analysis.
Nest-quality manipulation (experiment 3).—The basic arrangement for manipulation of the nest insulation was similar to nest material choice tests described above, but with the difference that only 1 grass tussock of either fresh (Group Fresh) or dried (Group Dry) fescue was presented to the animals for nest building. In this experiment we investigated how nest insulation depends on the moisture content and nest properties (wall thickness and nest mass) of the assembled nests. On the basis of the previous 2-way choice tests and on our earlier observations, 1 grass tussock of known volume, mass, and composition (moist Festuca or dried Festucd) was presented to the animals for nest building between 0800 and 0700 h (23 h). Eleven hours of light was sufficient for the animals to complete a nest of either material. Each animal was allowed to construct 1 “dry” and 1 “fresh” nest. Upon completion of the nests the following morning we removed the nest boxes with the nests and provided new material and a new nest box.
To determine the insulation value in relation to grass condition and moisture content we manipulated the moisture content of the nests in a sequence of 4 phases. In each phase the quality of nest insulation was measured. The 4 phases correspond to different levels of moisture content of the nest material on a dry-weight basis. First, we determined the difference in weight between fresh and completely dried nest material in a pilot study (not included in the present data set). When fresh fescue was dried in a laboratory drier until mass remained constant (i.e., essentially 100% solid), one-half of the original weight was lost, indicating that fresh material contained about 50% water.
Group Fresh nests (n = 8) were built from fresh, humid Festuca, and the insulation was measured in all 4 phases. In each phase the moisture content of the otherwise intact nests was set to a certain level. In phase 1 the nest contained 50% water (water content is 100% on a dry-weight basis). In phase 2 the nest contained 25% water (water content is 50% on a dry-weight basis). In phase 3 the nest contained 0% water (water content is 0% on a dry-weight basis), and in phase 4 the nest again contained 25% water (water content is 50% on a dry-weight basis) by adding the appropriate amount of water with a water sprayer to the dried nest. Group Dried nests (n = 8) were built from dried Festuca, and the insulation was measured only twice corresponding to phases 3 and 4 of Group Fresh.
Finally, nests were removed from the boxes, and nest mass (g) was measured on a dry-weight basis. The thickness of the nest wall and height of the nest also were measured (mm) at 5 randomly chosen points with a ruler, and the averaged value for each nest was used for further analysis. Additionally, lengths and widths of the slightly elliptical nests were measured.
The design of the experimental device followed the method of Redman et al. (1999). Glass bottles (150 ml) filled with hot (50°C ± 1 °C) water were inserted into each squirrel nest kept at room temperature (25°C ± 2°C). These bottles represented the animals within the nest, and the slower their temperature decreased the better the insulation was because thermal insulation refers to the decrease in rate of heat transfer. Each bottle was closed with a rubber lid through which a temperature-sensitive resistor (thermistor) was inserted. The temperature of the water bottle within the nest (Tb) and room temperature (Ta) were measured by thermistors. The sensors were previously calibrated to the nearest 0.1 °C against a precision quicksilver thermometer in a water bath between 0°C and 40°C. The signals from the thermistors were received with a model multimeter (M-4640A; Metex Instruments, Seoul, South Korea) via a channel multiplexer controlled and recorded by a personal computer. Finally, the multimeter was interfaced to a computerized system of data acquisition that recorded at 2-min intervals. Data recording lasted 5 h, long enough for the bottles to cool to room temperature.
The cooling constant (a; h−1) and insulation (I; °C/watt [°C/ W]) were calculated following the approach of Nicol and Andersen (2007). The relationship between time and cooling rate is best expressed by a negative exponential equation, where a was estimated by an iterative nonlinear regression procedure: where Tb – Ta is the difference between the temperature of water in the bottle (Tb) and the room temperature (Ta), b is the initial value of temperature difference, and ε refers to the residual variability (error), which has a normal distribution with 0 expected value. Based on the specific heat capacity of water (4.18 J g-1 °C−1) and bottle (0.84 J g−1 °C−1; mean ± SD mass: 102.66 ± 0.26 g) the thermal energy content of the 150 ml of heated water (50°C) equaled 35.65 kJ. For example, a 1°C drop in temperature over 1 min would equal 713 J°C, equivalent to 11.88 J/s or 11.88 W. At an initial temperature gradient of 25 °C between water bottle and ambient, the thermal conductance (C) would be 0.475 W/°C.
Statistical analyses—All statistics were performed using Statistica version 8.0 (StatSoft, Inc. 2008). For both the Bothriochloa-Festuca species preference experiment and the fresh-dry nest material choice experiment either a parametric paired Mest or a nonparametric Wilcoxon matched-pair test (Z) were used, depending on the distribution (normality) of data. The assumption of normality was tested with the Shapiro-Wilk test and quantile-qüantile (Q-Q) plots. In experiment 3, a mixed-model analysis of variance (ANOVA) was used to test the effects of moisture content in Festuca grass and nest material used for construction on nest insulation index.
To separate the effect of nest mass, nest height, nest length, nest width, and nest-wall thickness on nest insulation a best-subset regression model was used. The variance inflation factor was used to determine whether predictor variables were correlated (Hill and Lewicki 2006). To compare the insulation of nests made either from dry grass (phases 3 and 4; 2 cohorts) or fresh grass (phases 1–4; 4 cohorts), we used a nested ANOVA. The Bonferroni post hoc test was used for multiple comparisons following a significant ANOVA outcome. To examine relationships between insulation and both dry mass of nest and wall thickness Pearson correlation (r) was used. Summary data are reported as mean ± SD, unless stated otherwise. The criterion for statistical significance was P < 0.05.
Results
Nest material preference.—Squirrels (mean body mass: 172.5 ± 31.1 g) showed a preference for Festuca for nest material, both in terms of the composition of finished nests and time spent collecting nest material. The final nest contained significantly more Festuca (90% ± 5% of the nest material) than Bothriochloa (t4 = 18.82, P < 0.001). Squirrels also spent significantly more time in the vicinity of the Festuca tussock (median 82%, range 73-85%) than at the Bothriochloa tussock (Z = 2.02, n = 5, P < 0.05).
Quality of nest material—When subjects were given a choice between fresh and dry Festuca, they showed a preference for fresh Festuca (t7 = 4.24, P < 0.001); 97% ± 4% of nest material was fresh fescue. In addition, the animals spent more time in the vicinity of fresh tussocks (median 76%, range 67-85%) compared to dry ones (Z = 2.52, n = 8, P < 0.05).
Moisture content manipulation.—Both the quality of raw material (fresh versus dried: F1,35= 119.91, P < 0.0001) and moisture content (untreated, half-dried, dried, and half-moist, remoisturized: F4,35 = 21.54, P < 0.0001) influenced insulation significantly in a mixed-model ANOVA (Fig. 1). The differences in insulation (/) between untreated nests built from fresh, humid Festuca (phase 1) and posttreated nests built from fresh, humid Festuca (phases 2 and 3) were not significant (P > 0.05). However, I of phase 4 was significantly different (P < 0.0001) from phases 1, 2, and 3 built from fresh, humid Festuca. Mean thermal conductance values (in W/°C) of nests of cohort “fresh” were: Cphase_1 = 2.53 ± 0.19, Cphase_2 = 2.36 ± 0.15, Cphase_3 = 2.36 ± 0.13, and Cphase_4 = 2.94 ± 0.14. In the case of nests built from dried Festuca the difference in the insulation between untreated nests and half-moist, remoisturized nests was significant (P < 0.01; Fig. 1). Mean thermal conductance values (in W/°C) of nests of cohort “dry” were: Cphase _3 = 2.95 ± 0.21 and Cphase_4 = 3.31 ± 0.26.
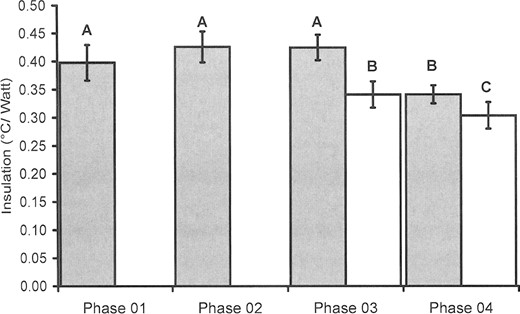
Mean (± SD) thermal insulation of nests constructed by European ground squirrels (Spermophilus citellus). Phases 1–4 refer to different moisture content (see text for details). Gray bars represent fresh nest materials (fescue) and white bars dried nest materials (fescue). Different letters above bars indicate significant differences (P < 0.01) between or among means.
Walls of the assembled nests made from fresh grass were significantly thicker (t14 = 4.36, P < 0.001) than the walls of the nests made from dried grass (Group Fresh = 4.28 ± 0.39 mm, Group Dried = 3.14 ± 0.63 mm; Table 1). Both the dry mass of the nest (r = 0.56, n = 16, P < 0.05) and the wall thickness (r = 0.80, n = 16, P < 0.0001) correlated with the insulation significantly. According to a best-possible-subset regression model, where nest mass, wall thickness, nest height, nest length, and nest width were entered as independent continuous predictors, and nest material (fresh or dry) as a categorical predictor, wall thickness and nest mass (F2,13 = 14.87, P < 0.001) explained 65% of the variation in nest insulation. Wall thickness by itself explained 61% of the variation in nest insulation (wall thickness: F1,16 = 16.16, P < 0.01; nest weight: F1,13 = 2.64, P > 0.05). The variance inflation factor was 0.8; therefore wall thickness and nest mass are considered uncorrected, reliable predictors.
Descriptive data (mean ± SD) for composition of fresh (n = 8) and dry (n = 8) nests constructed by European ground squirrels (Spermophilus citellus).
| Nest material | Wall thickness (cm) | Height (cm) | Length (cm) | Width (cm) | Dry mass (g) |
|---|---|---|---|---|---|
| Fresh | 4.28 ± 0.39 | 3.86 ± 0.41 | 19.63 ± 0.52 | 14.56 ± 0.82 | 19.25 ± 3.54 |
| Dry | 3.14 ± 0.63 | 3.48 ± 1.12 | 18.00 ± 2.14 | 14.06 ± 1.15 | 13.75 ± 2.71 |
| Nest material | Wall thickness (cm) | Height (cm) | Length (cm) | Width (cm) | Dry mass (g) |
|---|---|---|---|---|---|
| Fresh | 4.28 ± 0.39 | 3.86 ± 0.41 | 19.63 ± 0.52 | 14.56 ± 0.82 | 19.25 ± 3.54 |
| Dry | 3.14 ± 0.63 | 3.48 ± 1.12 | 18.00 ± 2.14 | 14.06 ± 1.15 | 13.75 ± 2.71 |
Descriptive data (mean ± SD) for composition of fresh (n = 8) and dry (n = 8) nests constructed by European ground squirrels (Spermophilus citellus).
| Nest material | Wall thickness (cm) | Height (cm) | Length (cm) | Width (cm) | Dry mass (g) |
|---|---|---|---|---|---|
| Fresh | 4.28 ± 0.39 | 3.86 ± 0.41 | 19.63 ± 0.52 | 14.56 ± 0.82 | 19.25 ± 3.54 |
| Dry | 3.14 ± 0.63 | 3.48 ± 1.12 | 18.00 ± 2.14 | 14.06 ± 1.15 | 13.75 ± 2.71 |
| Nest material | Wall thickness (cm) | Height (cm) | Length (cm) | Width (cm) | Dry mass (g) |
|---|---|---|---|---|---|
| Fresh | 4.28 ± 0.39 | 3.86 ± 0.41 | 19.63 ± 0.52 | 14.56 ± 0.82 | 19.25 ± 3.54 |
| Dry | 3.14 ± 0.63 | 3.48 ± 1.12 | 18.00 ± 2.14 | 14.06 ± 1.15 | 13.75 ± 2.71 |
Discussion
European ground squirrels showed a specific preference for fescue (F. pseudovina), one of the dominant grass species of grasslands inhabited by squirrels in Hungary. The preference was evident both in the behavior of animals during construction and in the composition of resulting nests in the laboratory choice tests.
Preference for fresh material was not expected. Thermal insulation refers to the capacity of materials to reduce heat transfer. Heat energy can be transferred by 4 main mecha-nisms: conduction, convection, radiation, and evaporation-condensation. According to thermal conductivity and evaporative heat transfer, higher water content of a material increases thermal conductivity and evaporation rate, and as a result reduces insulation. However, conductance was not affected by intracellular moisture content; that is, the insulation of nests made of fresh, humid Festuca did not increase with drying. Insulation is dependent on the thermo-physical properties (Bozikova and Hlavac 2005) and condition of the material. The insulation properties of feathers rank best, followed by grasses and then mosses (Hilton et al. 2004; Reid et al. 2002), but grasses are less affected by moisture than are feathers (Cramp 1977; Limona and Hoyo 1992). In contrast, the water spayed on the surface of the nest material decreased insulation significantly. The reduced negative effect on thermal conductivity of intracellular water within the leaves of xeromorph fescues (Festuca pseudovina, F. rupicola, and F. valesiaca) can be attributed to the external and internal morphological structures of their leaves (high ratio of air-filled sclerenchyma, constantly curled leaf blades, thick cuticles, pilosity, epistomic epidermis with a low stomata index, and recessed stomata under the epidermis). These characteristics help them resist dry and wet conditions (Kovács-Láng et al. 1995) and hence buffer the fluctuations in temperature and humidity within underground burrows (Meadows 1991). The negative effect of surface water on insulation probably was due to evaporative cooling.
Squirrels could build better-insulating nests from fresh grass than from dry grass, possibly because it provided a more flexible and treatable material, and they did not have to make a trade-off between fresh, humid and old, dry fescue. Finally, use of fresh grass resulted in better structure and thicker walls, which reduces air permeability of the nest and prevents penetration of cold air to the animal (Banks-Lee et al. 2004; Epps 1988; Van de Weerd et al. 1997).
The importance of nest architecture was supported by the result that wall thickness was positively correlated with insulation level, an effect that also was observed for nests of Microtus agrestis (Redman 1999). Nest insulation is not necessarily correlated with nest size (Ellison 1995), but our results agreed with those of Redman (1999) that wall thickness appears to be a better predictor than nest size for the insulation properties of a nest.
Nests should provide insulation for animals during stressful climatic periods. European ground squirrels construct under-ground nests below the freezing line, at a depth between 0.5 and 1.0 m (Hut and Scharff 1998; Ruzič 1978). The preference of fresh fescue grass in nest construction can be explained by the good insulation quality, which is a valuable feature in the active season when the temperature gradient between body and environment is great, but also during the normothermic periods of hibernation when the temperature gradient between body and environment is greatest (particularly prior to final emergence).
Dietary studies of European ground squirrels (Koósz 2002), European rabbits (Oryctolagus cuniculus), and European hares (Lepus europaeus—Katona et al. 2004) have shown the importance of xeromorph fescues; however, those studies emphasized only the importance of these plants in the diet. F. pseudovina is a common, but not the most frequent, element in the diet of squirrels (C. I. Gedeon, pers. obs.). Its use also as nest material (at least in the case of burrowing, nesting animals) was indicated by our results, but because it resists either dry or wet conditions, it might serve as a useful food item at the end of hibernation before emergence from burrows. To survive adverse environmental periods the quality of nesting material can be as important as food items, and fescue might serve both functions. This notion requires additional field and laboratory study. Experimental evaluation of specific abiotic (Váczi et al. 2006; Wagner and Drickamer 2004) and biotic (Kis et al. 1998) components could reveal characteristics of the habitat that are key to survival and thus provide valuable information for conservation efforts. Our results suggest that squirrels may rely on dominant, abundant, and readily available xeromorph fescues of dry meadows because of their importance to nest building.
Acknowledgments
We thank L. Porupcsanszki, Head of the Dunakeszi Airport, and the Duna-Ipoly National Park for permitting us to trap protected animals in the airport area. We thank M. Dombos, A. Horváth, O. Váczi, B. Koósz, P. Sandy, and A. Kövér for suggestions regarding the manuscript. In addition, we express our thanks to the anonymous reviewers for their useful and helpful comments, and in particular to L. C. Drickamer for thoroughly editing the text.
Literature Cited
Author notes
Associate Editor was Fritz Geiser.

{kind=link}