





Abstract
Perspectives on the importance of natural salt licks to ungulates have been broadened beyond the role of providing sodium. This study examined the chemical compositions of wet and dry licks in north-central British Columbia, and defined the benefits of licks to elk (Cervus elaphus), moose (Alces alces), Stone's sheep (Ovis dalli stonei), and mountain goats (Oreamnos americanus). We analyzed soils for buffering compounds (carbonates) and estimated available elements from extractions with solutions at a low, digestive-tract pH. Even though the 2 types of licks are visually distinct, with different concentrations of chemical components, they serve similar functions as concentrated sources of sodium, carbonates, magnesium, and sulfate. Sodium and sulfate concentrations were typically higher at both wet and dry licks than control sites. Carbonates and magnesium were elevated in soils from dry licks and magnesium also was high in inflow waters to wet licks. We estimated elemental intake by ungulates from the composition of forage samples. Forages used by all 4 ungulate species were too low in sodium to meet requirements. Spring and summer forages contained high potassium levels. Licks in our study, therefore, provide ungulates with supplemental sources of sodium that are particularly beneficial to offset increasing demands during lactation and with carbonates to help stabilize rumen pH after forage changes in spring. Supplemental sources of magnesium may be actively sought by ungulates when high levels of dietary potassium affect absorption. To assess the importance of clay in soils ingested at licks, we determined the clay mineral types at licks and compared the buffering capacity of clay-filled fecal material collected at licks with fecal material collected away from licks. Further studies are needed to define the roles of clay in improving forage palatability and digestibility.
Areas where ungulates actively ingest soil have commonly been referred to as salt licks. This name implies that licks serve a universal role of supplementing diets with sodium. Sodium is the dominant cation in the body fluids of terrestrial animals and the potential exists, especially for herbivores, to become sodium-deficient (Robbins 1993). Wild ungulates have a strong attraction to sodium salts (Dalke et al. 1965; Fraser and Reardon 1980; Stockstad 1950) and many studies have reported elevated concentrations of sodium in lick material (Fraser and Reardon 1980; Fraser et al. 1980; Hebert and Cowan 1971; Kennedy et al. 1995; Knight and Mudge 1967; Reisenhoover and Peterson 1986; Tankersley 1984; Tankersley and Gasaway 1983; Tracy and McNaughton 1995; Weeks and Kirkpatrick 1976). However, the supposition that the use of licks is attributable to a universal function is refuted by results from other analyses of lick soils showing low sodium contents and the presence of other important elements (Chamberlin et al. 1977; Coates et al. 1990; Dormaar and Walker 1996; Heard and Williams 1990). Given this variation in chemical composition, licks may serve multiple functions for different species and sexes at different times of the year (Kreulen 1985). For example, during late gestation and early lactation, more female white-tailed deer (Odocoileus virginianus) used mineral licks with higher concentrations of magnesium, calcium, and phosphorous than salt licks (Atwood and Weeks 2003). Elements other than sodium such as magnesium (Heimer 1988), buffering compounds such as carbonates (Kreulen 1985), and binding agents such as clays (Klaus and Schmid 1998) suggest multiple reasons for the use of licks by ungulates.
Deficiencies in macro- and trace elements are not necessarily a result of limited dietary intake, but rather of digestive disorders associated with spring forage change. The attendance at licks by many ungulate species peaks in the spring (moose [Alces alces]—Tankersley and Gasaway 1983; elk [Cervus elaphus]—Ayotte 2004; Williams 1962; Dall's sheep [Ovis dalli dalli]—Heimer 1973; Tankersley 1984; and mountain goats [Oreamnos americanus]—Singer 1978). This period coincides with the increased physiological demands of lactation, growth, or weight regain, which can be aggravated by electrolyte loss related to the stress of abrupt changes in forage chemistry. One source of physiological stress associated with adapting from highly fibrous winter diets to lush spring plant growth results from a decrease in fiber and an increase in readily fermentable carbohydrates and proteins that alter rumen pH and impair proper microbial function. Ruminants exposed to sudden drops in dietary fiber produce less saliva, which is high in bicarbonates (Kreulen 1985). With less saliva, the buffering capacity of the rumen is reduced (Church 1975) and can lead to a drop in pH below optimal conditions for rumen microbes, creating various intestinal ailments that reduce appetite and weight gain (Klaus and Schmid 1998; Kreulen 1985).
Physiological stress also can occur when extremely high potassium levels reduce the absorption and retention of other elements (Klaus and Schmid 1998; Kreulen 1985). Early-season vegetation contains concentrations of potassium that elevate the osmotic pressure of the digestive tract, interfere with fecal water absorption, and lead to potentially harmful electrolyte losses (Klaus and Schmid 1998; Kreulen 1985; Weeks and Kirkpatrick 1976). The observed wet, unformed feces from ungulates in spring, therefore, imply a loss of physiologically important elements (Hebert and Cowan 1971; Heimer 1988; Jones and Hanson 1985).
Supplemental sources of carbonates during the transition to spring forage can improve the buffering capacity of the rumen when saliva secretion is compromised (Kreulen 1985). Clay minerals, which also enhance buffering capacity, are composed of sheets of aluminium-silicate crystals, often with high cation exchange capacities (CECs—Cairns-Smith and Hartman 1986). Within the inorganic fraction of soil, CEC is essentially due to the contribution by clay minerals, because there are very few cation exchange sites in the sand and silt fractions (Brady and Weil 1999). During spring forage change, specific clay minerals can be associated with the adsorption of potassium and short-chain fatty acids to cation exchange sites, helping to adjust and maintain proper osmotic balance and pH of the rumen (Klaus and Schmid 1998; Kreulen 1985). Some types of absorptive clays also are able to bind secondary plant compounds such as tannins (Johns and Duquette 1991) and improve digestibility and feed conversion in domestic animals (Smith 1992).
Studies on licks tend to focus on a single ungulate species using 1 type of lick (i.e., wet licks or dry licks). Wet licks are associated with apparent groundwater springs. Dry licks usually occur along streams or riverbeds, where unweathered deposits of soluble elements have concentrated above less-impervious layers, and become exposed by erosion. Attempts to compare the chemical composition of lick material across different types of licks in the context of the role of licks to different ungulate species are made difficult because of different methods employed in soil sampling and analytical procedures (Klaus and Schmid 1998). Lack of controls and overall low sample sizes precluded 8 of the 33 studies reviewed by Klaus and Schmid (1998) from using statistical tests to examine differences in the composition of lick soils. To represent available versus selected resources, control sites should remain in the same geological formation, but avoid seepage influences from the lick (Kennedy et al. 1995). To sample highversus low-use areas, spatial use of licks by ungulates should be documented before soil sampling (Ayotte 2004).
Critical to comparing the results from past research on licks is consistency in the methods used for chemical analyses. Klaus and Schmid (1998) summarized the common extraction methods as extracting the water-soluble fraction; extracting the water-soluble fraction and the cations adsorbed to colloids with ammonium acetate; and measuring the total elemental content of the soil with X-ray fluorescent analysis. Each method extracts a specific part of the sample, such that results from 2 different extraction methods on the same sample are not comparable. Many pretreatments of soil material can destroy and prevent detection of potentially important clay minerals (Kabata-Pendias and Pendias 1992). Very few studies have selected a pH for extraction solutions that is similar to the pH of absorption sites in the digestive tract (e.g., Mahaney et al. 1997).
The objective of this study was to determine the differences in chemical composition of lick soils within and among licks used by 4 ungulate species. It was part of a larger project that documented attendance patterns and licking intensity by elk, moose, Stone's sheep (Ovis dalli stonei), and mountain goats during spring and summer (Ayotte 2004). In northern British Columbia, Canada, wet licks are frequented by moose and elk, whereas Stone's sheep and mountain goats ingest soil at dry licks. We used consistent soil sampling and methods of analysis across multiple wet and dry licks in 1 watershed. We incorporated the compositions of lick soil, important forages, and fecal samples (comparing the loss of electrolytes between animals that had recently ingested lick material and those that had not) with a review of elemental requirements. We hypothesized that if forage content of specific elements was less than requirements, and if those elements were in greater concentration in lick soils than control sites, then those elements were actively sought by ungulates using licks. We predicted that concentrations of carbonates and clays, as buffering and binding compounds that help accommodate spring green-up and plant defensive compounds, would be higher in lick soils than control sites. Because the ungulate species in our study all experience spring forage change in the watershed, and the females of each species have increased demands associated with lactation, we hypothesized that even though wet and dry licks are visually distinct and used by different species, the relative chemical compositions would not differ between lick types.
Materials and Methods
Study site.—Samples of lick soil, forage species, and ungulate feces were collected in the Tuchodi watershed (58°10′N. 124°30′W) of Northern Rocky Mountains Provincial Park in north-central British Columbia, Canada. The study area extends approximately 55 km along the Tuchodi River system. Vegetation in the watershed is dominated by white spruce (Picea glauca) and black spruce (P. mariana) at low elevations; by black spruce, willow (Salix), and birch (Betula glandulosa) in the subalpine zone; and by alpine tundra. Six licks were chosen based on historic ungulate use, adequate observation points, and access by researchers. The 3 wet licks (Childers, Dead Dog, and Grizzly) were treeless areas of deep mud with a history of heavy use by moose and elk as indicated by the surrounding network of deeply eroded trails. The 3 dry licks (Lac-a-nookie, Standard, and Upriver) used by Stone's sheep and mountain goats were located along river cut-banks, near valley bottom, and in close proximity to steep escape terrain.
Soil collection and analyses.—Locations for soil sampling within a lick area were distinguished by the level of use (high, with clustering of animals, and low, with few or no animals). Distinctions between high- and low-use sites were based on ∼600 h of behavioral observations on elk and moose at 2 of the 3 wet licks and on Stone's sheep and mountain goats at 3 dry licks, from 17 May to 11 August 2002 (Ayotte 2004). All animal locations were plotted on a 4 × 4-m grid map of the lick area. The 4 × 4-m dimensions reflect the accuracy of an observer in nearby blinds recording animal locations. The boundaries of the licks were determined by the edge of exposed soil. Soil samples were collected between the end of July and early August to allow sufficient observations to establish the locations of high-use sites within the lick areas and to avoid disturbing animals at licks during peak use. We collected samples from 9–11 sites in each of high-use, low-use, and control areas (Klaus and Schmid 1998) at each lick; all sampling sites were 4 × 4 m. Control sites were exposed soil with no signs of use, sampled 10 m beyond the edge of the lick area, with sites spread evenly around the perimeter of the lick unless possible seepage influences were suspected. Soil was collected from a 3rd wet lick (Grizzly) without previous observations of animal use and analyzed relative to lick and control sites.
Within each high-use, low-use, and control site, composite samples of 12 subsamples were collected and pooled to ensure a consistent sampling intensity among sites. However, several high-use sites at dry licks were sampled from isolated excavations smaller than 4 × 4 m. All composite samples from all sites weighed approximately 300 g. A total of 178 soil samples were collected and analyzed.
Soil samples were air-dried in paper bags (Stockstad et al. 1953), ground lightly (Dormaar and Walker 1996) with a glass rolling pin, and passed through a 2-mm sieve (Dormaar and Walker 1996; Stockstad et al. 1953) to homogenize the sample before chemical analyses. We used subsamples of the fine soil fractions to determine pH; carbonate equivalent; CEC; available macroelements (Na, Ca, Mg, K, P, and S) and trace elements (Fe, Mn, Cu, and Zn) at pH 2.5; total carbon and nitrogen; and clay fraction and clay mineral type.
The pH of each soil sample was determined in 0.01 M CaCl2 suspensions equilibrated overnight (Peech 1965). The calcium carbonate equivalent was determined by an acid-neutralization procedure (United States Department of Agriculture 1954) as an estimate of total carbonates (CO3; British Columbia Ministry of Forests Analytical Research Lab, Victoria, British Columbia, Canada). We quantified soil CEC as centimoles of positive charge per kilogram of soil (cmol(+)/kg; equivalent to milliequivalents per 100 g) by extracting samples with 1 N ammonium-acetate adjusted to pH 7 (Kalra and Maynard 1991) to reflect the pH of the rumen (5.5–7—Church 1975). We specifically chose a pH 2.5 extraction solution to estimate concentrations of elements that become available to the animal as the lick soil passes through the abomassum (pH ∼ 2.5) just before the major sites of elemental absorption in the small intestine (Church 1975). All soil samples were analyzed (British Columbia Ministry of Forests Analytical Research Lab) for cations (Na, Mg, Ca, K, P, Mn, Zn, Cu, Fe, Co, and B) using the Mehlich III procedure (Sen Tran and Simard 1993) because the pH of the extractant is very close to 2.5. Sulfur in the form of sulfate (SO4) was the only anion extracted using a 500-ppm phosphorus method (Kowalenko 1993) with extraction solutions adjusted to pH 2.5. Total C and N were determined by an NCS autoanalyzer (NA1500, Fisons, Milan, Italy).
We used nested analysis of variance (ANOVA) with use level (high use, low use, and control) nested within lick (Lac-a-nookie, Standard, Upriver, Childers, Dead Dog, and Grizzly) to test for significant differences in the concentrations of each element extracted at pH 2.5, carbonates, sulfate, CEC, and pH among high-use, low-use, and control sites at each lick. Data were transformed with log or Box–Cox power transformations (Box and Cox 1964) where appropriate to meet assumptions of normality and homogeneity of variance (Levene's test—Zar 1999). Bonferroni tests were used for all post hoc analyses to adjust for the high number of contrasts (Sokal and Rohlf 1995). These analyses were only conducted for the 3 dry licks and 2 wet licks for which previous animal observations had quantified use level. At Grizzly lick where high and low animal use was not differentiated, we used simple 1-way ANOVA to compare use and control sites.
Clay separations and identification of clay mineral types were conducted on a small subsample of soils. Three high-use and 3 control samples from 1 wet lick (Childers) and 1 dry lick (Lac-a-nookie) were separated into sand, silt, and clay fractions after pretreating samples to remove organic matter and carbonates (Kalra and Maynard 1991); results were then averaged by fraction. Slides of isolated clays were scanned with an X-ray diffractometer (D8-GADDS; Bruker, Madison, Wisconsin) using methods adapted from Whittig (1965) to determine mineral composition and the presence or absence of expandable clays that have high CEC. The expanding nature of these clays benefit the ruminant by binding and removing acidic compounds from the digestive tract.
Water collection and analysis.—Water samples from 2 wet licks (Childers and Dead Dog) were collected. We obtained 3 samples from the inflow (apparent groundwater spring), the outflow, and from control sites (the closest body of water outside the lick) for a total of 9 samples from each lick. A few drops of concentrated nitric acid were added to samples to preserve trace elements in solution for later analysis. The cations in each water sample (Na, Mg, K, Ca, P, Fe, Mn, Cu, Zn, Co, and Mo) were analyzed with inductively coupled plasma–atomic emission spectroscopy (PS 1000UV, Leeman Labs, Inc., Lowell, Massachusetts).
We used nested ANOVA with source location nested within lick to test for significant differences in concentrations of each element among inflow, outflow, and control water samples. Tukey's test was used for post hoc comparisons (Sokal and Rohlf 1995).
Forage collections and analyses.—Three collections of forage were made for each ungulate species: forage for moose and elk was collected in early June, early July, and late July; and forage for Stone's sheep and mountain goats was collected in early June, mid-July, and early August. We attempted to collect 6 species of forage used by each ungulate species at 3 different elevation zones in the Tuchodi watershed during each period. These plant species were identified previously as primary dietary components in several local and regional studies. Grasses and herbs were generally collected as whole plants, whereas leaves and twigs were sampled from browse species.
Primary forage plants of elk have been reported to be Elymus, Bromus, Vicea americana, Vibernum edule, Rosa acicularis, and Populus balsamifera (Peck 1987). Composition of elk diets changes from 90% grasses, 7% herbs, and 3% shrubs in spring to 17% grasses, 50% herbs, and 33% shrubs in summer (Peck 1987). Dominant forage plants for moose are Cornus canadensis, Epilobium, Alnus, Salix, Betula papyrifera, and Populus balsamifera (Cushwa and Coady 1976; Singleton 1976). We did not have data available to distinguish between percentage composition of spring and summer diets of moose, and therefore we assumed the composition of the diets in both seasons to be 5% herbs, 20% Alnus/Cornus/Populus, and 75% Salix/Betula (Cushwa and Coady 1976; Singleton 1976). Principal forage species of Stone's sheep include grasses and herbs (Agropyron, Elymus, Poa, Hordeae, Oxytropis, and Zygadenus), with 80% grasses and 20% herbs in spring diets and 60% grasses and 40% herbs in summer diets (Luckhurst 1973; Seip 1983). Mountain goats consume grasses (Agropyron and Festuca altaica), herbs (Lupinus, Epilobium, and Artemesia), and sedges (Carex). Again, because distinctions have not been reported between spring and summer diets of mountain goats, we assumed both to be 5% grasses, 75% herbs, and 20% Carex (Hjeljord 1973).
We selected forage collection areas in each elevation zone using signs of recent ungulate presence (grazing or browsing, pellets, and tracks), mainly on south- and west-facing slopes. The lowest elevation zone (800–959 m) for collection of elk and moose forage was in the valley bottom near the wet licks, in stands of aspen and balsam poplar (Populus) within the boreal white spruce and black spruce bio-geoclimatic zone (Meidinger and Pojar 1991). The mid-elevation zone (1,000–1,150 m) was in young shrub and sapling communities in the boreal white spruce and black spruce zone, and the high zone (1,250–1,400 m) was in open poplar stands and upland shrub communities in the spruce, willow, and birch zone (Peck 1987). Collection areas for Stone's sheep and mountain goat forage began close to the valley bottom, in plant communities similar to those for elk and moose forage collections, but at slightly higher elevations and often near rock outcrops, with the highest zone (1,400–1,650 m) in alpine plant communities. Each forage sample from an elevation zone consisted of ∼ 10 randomly chosen plants. Each sample weighed ∼250 g to ensure a dry weight of 25 g for analysis (Ohlson and Staaland 2001). Plant samples were air-dried in the field and later oven-dried for 72 h at 40°C before being ground and sieved with a Wiley mill (Thomas-Wiley, Swedesboro, New Jersey) using a 0.25-mm sieve screen (Staaland and White 2001). We prepared plant samples for elemental analysis by microwave digestion using nitric acid and hydrogen peroxide (Kalra and Maynard 1991). Plant extracts were analyzed for macro elements (Na, Mg, K, Ca, and P) and trace elements (Fe, Mn, Cu, Zn, Co, and Mo) using an inductively coupled plasma–atomic emission spectrometer (PS 1000UV, Leeman Labs).
To examine the elemental variability in the diet of each ungulate species over time, we averaged data from the forage species within functional groups (e.g., grasses, herbs, and shrubs) by collection period and then weighted them by their proportion in the hypothetical diet to obtain 1 dietary estimate for each elevation zone. Because 1-way ANOVA indicated that there were no effects of elevation zone (all P > 0.78), we considered samples at each elevation as replicates. We then used 1-way ANOVA with collection period as the independent variable to test for significant differences in the concentrations of each dietary element among collection periods; Tukey's test was used for post hoc analyses (Zar 1999). During the early-June collection period, browse plants consumed by elk and moose were only available (and therefore collected) at low elevations, resulting in a single estimate of diet compared to 3 (low, mid-, and high elevations) for other collection periods. Accordingly, we used 1-way ANOVAs (Zar 1999) to determine significant differences in elemental concentrations only between the 2nd (early July) and 3rd (late July) collection periods.
Fecal sample collections and analyses.—Fecal samples were collected at all lick areas (lick fecal samples) and from distances at least 2 km from licks (nonlick fecal samples). At lick areas we collected fecal samples that contained a noticeable amount of inorganic material (typically white or gray-colored in fresh samples). Only fecal samples that appeared organic were collected away from licks and grouped as nonlick fecal samples. Each fecal sample consisted of material from 1 pellet group. Elk and moose pellets were distinguishable, but Stone's sheep and mountain goat pellets were not and so were grouped together.
Because the CEC of digesta in the gastrointestinal tract of ungulates that have ingested soil is made up of both inorganic and organic contributions, analyses were conducted on both inorganic and organic components in the feces. Fecal samples were air-dried in the field and later oven-dried for 72 h at 40°C before being ground and sieved (Wiley mill with 0.25-mm sieve). Five grams of each sample were left overnight in 1 N ammonium acetate at pH 7 solutions to extract cations and determine CEC with the same procedures used on soil samples. The extractions were analyzed for macroelements (Na, Mg, K, and Ca) and trace elements (Fe, Mn, Cu, Zn, Co, and Mo) by inductively coupled plasma–atomic emission spectroscopy. We tested for differences in the concentrations of cations between lick and nonlick fecal material using 1-way ANOVA with equal or unequal variances as appropriate (StataCorp 2005).
To quantify the amount of organic matter in each fecal sample, a few milligrams of the remaining unextracted sample were reground with liquid nitrogen (rather than a mechanical grinder to conserve material because samples were small), and analyzed for total carbon with an elemental analyzer. The percentage organic matter was calculated from the amount of total carbon (organic matter = 1.72 × percent carbon—Brady and Weil 1999) and used to estimate its contribution to the total CEC of the sample. The contribution of inorganic clay to an animal's buffering capacity was determined by the difference between total and organic CEC.
All values for soil, water, forage, and fecal samples are presented as untransformed means ± 1 SE unless otherwise noted. We used Stata 9 (StataCorp 2005) and STATISTICA 6.1 (StatSoft Inc. 2002) software for all tests; significant differences were assumed at α = 0.05.
Results
Concentrations of elements, CEC, and pH in lick soils were all significantly different among licks (all P < 0.001) except for potassium concentration (P = 0.229). Concentrations also were different among use levels (high-use, low-use, and control sites; all P < 0.001). Our results show that geochemistry varied between licks of the same type (e.g., wet or dry) and among sites at the same lick.
Wet licks.—There were no consistent patterns in the mean concentrations of soil components among the 3 wet licks. For example, Childers lick soils contained twice the concentration of calcium, 10 times the concentration of sulfate, 6 times the CEC, and one-third the copper concentration of Dead Dog and Grizzly licks (Fig. 1). Sodium concentrations were higher in lick soils, although not significantly above control sites. Carbonate content also tended to be higher, whereas total carbon and nitrogen contents and CEC were typically lower than for controls (Fig. 1). Sample sizes, however, were too small to detect significant differences at all licks.
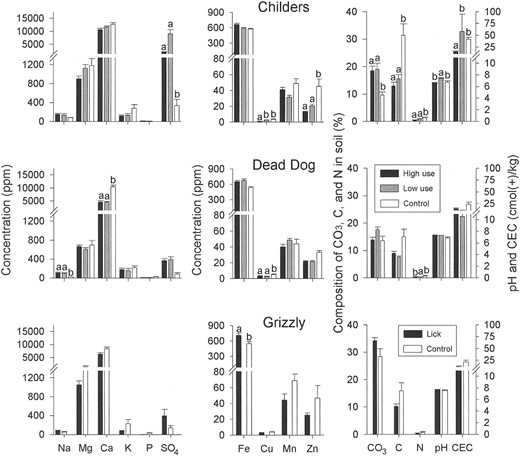
Concentrations (mean + SE) of soil components in samples collected from high-use, low-use, and control sites (n = 9–10 for each type of site) at 3 wet licks in the Tuchodi watershed, British Columbia, Canada, 2002. All units are in ppm except for CO3 (%), C (%), N (%), cation exchange capacity (CEC; cmol(+)/kg), and pH. Note that for Grizzly site there were no data on use, so comparison is for lick and control only. Soil components with letters above bars were significantly different between use sites within licks via nested analysis of variance; values sharing the same letter were not different according to Bonferroni contrasts.
Clay content of high-use soils from Childers lick was similar to controls (high-use: 63.2% ± 3.0%, n = 3; controls: 67.2% ± 2.5%, n = 3; P = 0.370). X-ray analyses of isolated clay fractions showed no differences in the type of clay mineral; both high-use and control samples contained mica, quartz, and chlorite. No expandable clays (high CEC) were present in any of these isolated clay fractions.
Elemental composition of water samples varied among licks. For example, sodium concentrations in water entering Dead Dog lick were 5 times higher than those at Childers lick (Fig. 2). Within licks, sodium, magnesium, calcium, and molybdenum concentrations always were greater in inflows than in controls (Fig. 2). At Childers lick, the inflow and outflow concentrations of these elements were equivalent. In contrast to Childers lick, outflows and controls at Dead Dog lick were not different. High levels of potassium were observed only in inflow water to Dead Dog lick.
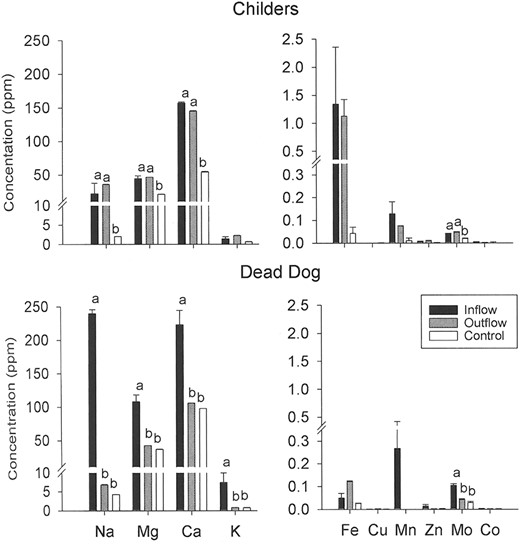
Elemental composition (mean + SE) of water samples collected at 2 wet licks (n = 3) for each inflow, outflow, and a control in the Tuchodi watershed, British Columbia, Canada, 2003. Elements with letters above bars were significantly different as shown by nested analysis of variance; within soil components, bars sharing the same letter were not different according to Tukey's multiple range test.
Dry licks.—Concentrations of elements and compounds were relatively similar across all 3 dry licks except for sodium, zinc, carbonates, and CEC (Fig. 3). Soils from Lac-a-nookie and Standard licks, which are both steep gravel bluffs at the same elevation, contained carbonates that were 10% higher, and concentrations of zinc and CEC that were one-third lower than at the physically different Upriver lick. Sodium concentrations in soils from Lac-a-nookie, however, were more similar to those from Upriver lick and were nearly twice the levels at Standard lick.
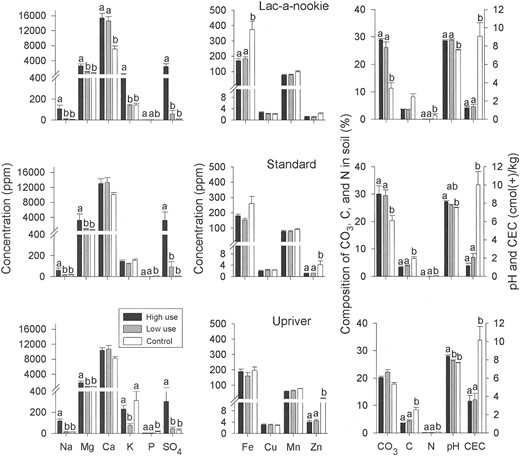
Concentrations (mean + SE) of soil components in samples collected from high-use, low-use, and control sites (n = 9–11 for each type of site) at 3 dry licks in the Tuchodi watershed, British Columbia, Canada, 2002. All units are in ppm except for CO3 (%), C (%), N (%), cation exchange capacity (CEC; cmol(+)/kg), and pH. Soil components with letters above bars were significantly different between use sites within licks as shown by nested analysis of variance; values sharing the same letter were not different according to Bonferroni contrasts.
The chemical composition of soil from dry licks differed from that of wet licks. In contrast to wet licks, concentrations of sodium, magnesium, and sulfate, as well as pH, always were highest at high-use sites. Phosphorus and nitrogen levels in high- and low-use sites were lower than in control sites. Similar to wet licks, total carbon, nitrogen, and CEC of soils from the dry licks tended to be lower than from controls.
High-use soils analyzed from Lac-a-nookie lick contained more than twice the amount of clay as control samples (high-use: 49.7% ± 0.7%, n = 3; control: 20.8% ± 8.4%, n = 3; P = 0.026). There were no differences in the types of clay minerals present in the isolated clay fractions of high-use samples compared to controls. Quartz, mica, and chlorite were present in all samples, and no expandable clays with high CEC were found in any samples.
Forage.—Statistical testing of the seasonal elemental variation in forage in elk and moose diets was limited by small sample sizes during spring forage collections in early June. The diets of elk, however, were high in potassium and sodium and low in iron during the spring forage collection period (Fig. 4). Levels of phosphorous, manganese, and molybdenum were significantly higher (all P ≤ 0.04) in elk forage in early July compared to late July. No significant differences in elemental concentrations were observed in moose diets across the summer. Sheep diets were elevated in calcium and molybdenum in early June and low in cobalt in early August (Fig. 4; all P ≤ 0.01). Mountain goat diets were high in phosphorus in early June and were also low in cobalt in early August (Fig. 4; all P ≤ 0.02).
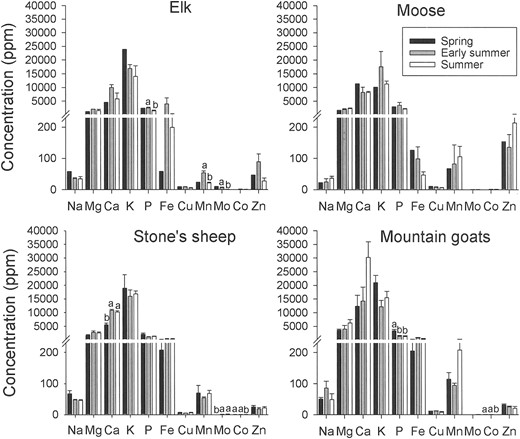
Elemental composition of ungulate diets based on important forage plants sampled in the Tuchodi watershed, British Columbia, Canada, during 3 collection periods: spring (early June), early summer (early and mid-July), and summer (late July and early August), 2002. Data are presented as mean + SE. Forage elements with letters above bars were significantly different between collection periods via analysis of variance (only early-summer and summer periods could be tested statistically for elk and moose because of missing samples); within elements, bars sharing the same letter were not different according to Tukey's multiple range test.
Fecal samples.—Only elk fecal samples from wet licks were analyzed. Samples from moose were too few to present in a comparison with nonlick samples. The inorganic content of feces was not significantly different between samples collected at wet licks and those collected at distances away from wet licks (at wet licks: 14.4% ± 2.8%, n = 6; away from wet licks: 17.3% ± 4.5%, n = 14; P = 0.68). In contrast, fecal samples that were collected at dry licks contained significantly more inorganic material than fecal samples from distances away from dry licks (at dry licks: 80.1% ± 2.4%, n = 22; away from dry licks: 15.5% ± 1.3%, n = 10; P < 0.001). The observed CEC (contributed to by both organic and inorganic fractions of fecal samples) of 17 of 21 samples was lower than the expected CEC contribution of organic material alone in organic soil types (297 cmol(+)/kg—Stevenson 1994).
The concentrations of sodium, calcium, and copper in elk feces collected at wet licks were significantly lower than feces collected away from licks (Fig. 5). Stone's sheep and mountain goat fecal samples collected at dry licks contained more magnesium, iron, molybdenum, and cobalt and less sodium, potassium, copper, and zinc than did nonlick fecal material (Fig. 5).
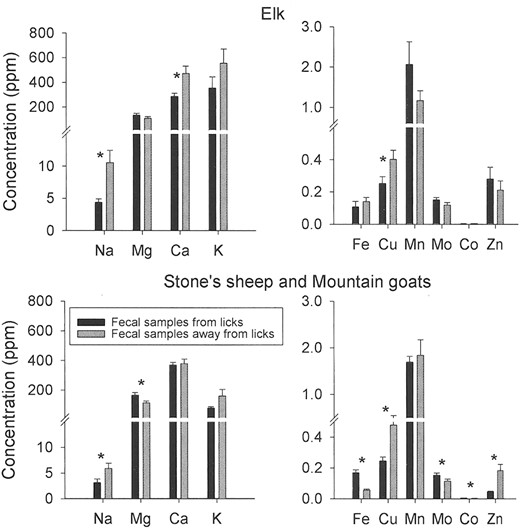
Cations extracted with ammonium acetate at pH 7 from fecal material (top: elk; bottom: Stone's sheep and mountain goats) collected at licks and from nonlick areas >2 km away from licks in the Tuchodi watershed, British Columbia, Canada (June–July, 2002 and 2003). Data are presented as mean + SE (elk: lick n = 15, nonlick n = 17; Stone's sheep and mountain goats: lick n = 32, nonlick n = 17). An asterisk (*) indicates significant difference between cation concentrations of fecal samples from licks and away from licks.
Discussion
The variation in composition of lick soils across previous studies has called into question a universal function of licks as salt licks, and suggested that different licks serve different roles (Kreulen 1985). Our results indicate that each lick serves multiple functions and that although there is variation between lick types and among sites within licks, there are similarities in function across all licks in the study area.
Buffering capacity.—Carbonate content and pH were higher (although not always significantly) in soils from all 6 licks (wet and dry) than from control sites. The elevated concentrations of carbonates in lick soils in the Tuchodi watershed would be important in buffering the rumen of ungulates during the transition each spring from fibrous winter forage to lush spring growth, as documented in other studies (e.g., Bechtold 1996; Fraser et al. 1980; Knight and Mudge 1967). Fluctuation in rumen pH is a critical factor in rumen microbiology, and salivary bicarbonates buffer against the rumen pH becoming too acidic (Church 1975). In both salivary bicarbonate and the inorganic carbonates found in soils, the hydrolyzing ability of carbonate ions contributes to the buffering capacity by neutralizing the acidity produced in the rumen (Brady and Weil 1999).
A drop in rumen pH below optimum levels (e.g., ≤4.3–5.3 for domestic sheep—Church 1975) may cause a number of ailments such as depressed forage intake, diarrhea, and reduced absorption of macro- and trace elements that are in solution in the digestive tract (Kreulen 1985). The effectiveness of sodium bicarbonate in alleviating acidosis and improving feed conversion and milk production in dairy cows on concentrate feed has been consistently noted in domestic ruminant research (Fisher and Mackay 1983). Susceptibility to the conditions of acidosis is also increased by poor body condition (Kreulen 1985). Therefore, concentrations of carbonates in exposed lick soils would help ungulates minimize acidosis after spring forage change, especially those individuals in poor condition after severe winters.
Ingesting clay soils from licks also would improve buffering capacity and adjust rumen pH. At the 1 dry lick that we analyzed, the clay content was higher than in control samples. At both types of licks, however, clay content of the lick soil was ≥50%. We were unable to confirm the presence of expandable clays in soils from either wet or dry licks, but sample sizes were very low. Other studies have noted that licks are associated with subsoils of high clay content (Weeks 1978) or clay-rich pans that hold water (Holdø et al. 2002), but the potential contribution of the clay components per se to the nutritional ecology of ungulates has not been addressed.
The inorganic fraction in fecal samples, which is reflective of soil ingestion, also can reflect improved buffering capacity. At dry licks in particular, the inorganic content of fecal samples averaged 80%. The cation exchange sites in inorganic fractions are largely due to clay particles that have greater surface area with negative charge than sand and silt (McBride 1994). Only 4 fecal samples in our analyses had total CEC higher than the expected CEC contributed to by organic matter, implying that in those samples the inorganic matter increased the samples' total CEC. This assessment of the contribution of clays may be underestimated, however, because of the relative contribution to the CEC by highly decomposed organic matter. Decomposition of organic matter and its effect on surface charge determine the contribution from organic matter to the overall CEC (Deng and Dixon 2002). The level of decomposition of the organic matter in fecal pellets is likely higher than that of the organic matter in the rumen, where the buffering role of clay is considered important (Klaus and Schmid 1998; Kreulen 1985). Therefore, clay ingested from lick soil would probably contribute to the CEC and buffering capacity in the rumen, particularly if the level of decomposition of rumen organic material was low. In addition, the CEC and buffering capacity of clays are less affected by the low-pH conditions of rumen acidosis than are those of organic matter (Brady and Weil 1999).
Ingested clay also has the potential to bind secondary plant compounds such as tannins that decrease forage digestibility (Johns and Duquette 1991; Smith 1992). This would be most useful to species such as moose and mountain goats that forage on a wide variety of plants and may consume significant amounts of browse and herbaceous forages that produce defensive compounds. Highest use of licks in mid-July by moose (Ayotte 2004) corresponds with high forage biomass and intake of tannins at this time. Similarly, use of lick soils containing clays could help to ameliorate the digestive ailments (i.e., soft, unformed diarrheic feces) frequently observed in mountain goats in midsummer.
Elemental contributions.—Across all licks, there were no elements that were consistently elevated in lick soils compared to control sites, but there were several that were common to each type of lick soil. The significance of these elements is illustrated by comparing soil and forage analyses with ungulate requirements. We used our forage analyses as estimates of general trends in the availability of elements in the diet, although we recognize that there are probably complex mechanisms allowing ungulates to balance their requirements of the 24 elements essential for growth and reproduction (Staaland and White 2001).
Sodium in soil was significantly elevated above controls at all 3 dry licks and 1 wet lick (Dead Dog lick). At that wet lick, the highest use areas also were associated with water inflows having the highest sodium concentrations measured (240 ppm). At the dry licks, where Stone's sheep and mountain goats were observed ingesting soil from isolated pockets, high-use sites contained significantly higher sodium than low-use sites and controls. Chemical differences between high- and low-use sites at wet licks were not detectable because of the effects of soil mixing, movement of water through the lick, and the absence of isolated pockets. Elevated concentrations of sodium in lick soils have been reported frequently (e.g., Bechtold 1996; Dormaar and Walker 1996; Jones and Hanson 1985).
Estimates of the sodium required by captive ungulates as a component of dry matter intake (600–1,600 ppm for maintenance and lactation—Church 1975; Lintzenich and Ward 1997; National Research Council 1996) are much greater than the average concentrations of sodium (<86 ppm) measured in the diets of the 4 ungulate species in our study area. Sodium demands are increased by 40% of predicted maintenance requirements during early lactation (Staaland et al. 1980), corresponding with periods of high use of wet licks by female elk (late June—Ayotte 2004). To minimize predator threats to young while traveling from alpine foraging habitats to licks on valley bottoms, visits to dry licks by female Stone's sheep and mountain goats may be delayed until later in the summer (early July and early August—Ayotte 2004). Sodium demands of antlerogenesis (136 ppm in white-tailed deer) are small compared to those of lactation (Hellgren and Pitts 1997), although the increased demand (especially during the latter half of the antlerogenesis period when growth is highest—Bubenik 1982) may precipitate sodium deficiency in some male cervids, stimulating lick use. The strong selective appetite for sodium shown by wild ungulates (Dalke et al. 1965; Fraser and Reardon 1980; Stockstad 1950) implies an adaptive response to sodium stress in which depleted sodium pools may be replaced by short periods of high intake (Staaland et al. 1980). The high sodium concentrations in dry lick soils (60–120 ppm) and in water entering wet licks (22–240 ppm) can provide an important source of sodium in a concentrated area. Assuming that fecal sodium can be used as an indicator of sodium balance (Holdø et al. 2002), the individuals using both wet and dry licks excreted less sodium, which is consistent with a sodium supplementation role of licks.
Sodium stress may be exacerbated by high levels of potassium in forage that inhibit the retention of sodium and other elements in solution in the fecal tract (Kreulen 1985). Staaland et al. (1980) found that lowest plasma sodium levels in reindeer (Rangifer tarandus) were associated with summer forage high in potassium and low in sodium. If the sodium-to-potassium ratio in saliva ranges below 1.4:1–1.6:1 (Staaland et al. 1980), sodium deficiencies are aggravated. The ratio of sodium to potassium in the diets of all ungulate species in our study was <0.004:l. Although not directly related to saliva production, this ratio implies an osmotic imbalance that supplemental sources of inorganic material from licks could alleviate. Atwood and Weeks (2002) noted that potassium-induced sodium deficiency compels animals to seek ancillary sources of sodium. Supplementing domestic sheep feed with calcium carbonate has been shown to elevate urinary potassium excretion (Yano et al. 1979), reducing the inhibitory effects on sodium. Some clay minerals also have been shown to alleviate potassium imbalances by stimulating potassium excretion (Brady and Weil 1999; Kreulen 1985). X-ray analyses on a larger number of soil samples from each lick are needed before ruling out the presence of these clay minerals and their potential importance to ungulate species using licks.
Sulfur in the form of sulfate was significantly concentrated in lick soil at all 3 dry licks and was almost significantly concentrated at 1 wet lick (Dead Dog, P = 0.052; Figs. 1 and 3). Concentrations of sulfate in lick materials have rarely been reported (Heard and Williams 1990). Sulfur comprises 4–5% of hair and wool and is an important element in rumen microbe metabolism (Church 1975). Sulfur deficiencies are associated with deficiencies in sulfur-containing amino acids (methionine and cysteine—Robbins 1993) that may cause decreased appetite and milk production, and poor pelage (Church 1975). We were unable to quantify the concentration of sulfate (an anion) in forage samples with inductively coupled plasma–atomic emission spectroscopy for cations to compare with concentrations in lick soil, and, therefore, could not determine if sulfate from licks might be meeting a nutritional deficiency. Given that the elevated levels of sulfate in lick soil may result from the dissolution of sodium sulfate minerals, however, ungulates could benefit from access to sulfate-containing soils when fulfilling sodium requirements at licks.
Magnesium was elevated in water flowing into wet licks (close to the high-use sites) and in soil at dry licks. Elevated concentrations of magnesium are common in lick soils across North America (e.g., Bechtold 1996; Heimer 1988; Jones and Hanson 1985). The inaccessible form of stored magnesium in the mammalian body (70% in bone—Robbins 1993) and the common occurrence of grass tetany (low serum magnesium) in domestic livestock (Heimer 1988) support the importance of a supplementary source of magnesium. In a comparative study of natural versus artificial licks, Atwood and Weeks (2002) reported that more white-tailed deer frequented natural licks where sodium levels were lower and magnesium and sulfate levels were both higher than at artificial licks. Although estimated levels of dietary magnesium for the ungulate species in our study (1,187–6,221 ppm) were close to meeting requirements (900–2,000 ppm for maintenance and reproduction— Lintzenich and Ward 1997; National Research Council 1996), other dietary properties of spring forage, particularly high potassium levels (Fig. 4), can reduce the absorption of magnesium, especially in lactating ungulates in the spring (Church 1975; Robbins 1993). Therefore, lick soil and water are important sources of magnesium during the transition to spring forage. Ingesting soil from licks is a fundamental component of the foraging strategies of many ungulates in north-central British Columbia. For these species that typically ingest very low amounts of soil incidental to their feeding (Beyer et al. 1994), soil ingestion at licks is a deliberate action to meet demands. Furthermore, this drive to visit licks is at the expense of time and energy spent traveling that could have been spent foraging (Klein and Thing 1989) and of associated predation risks (Hui 2004). The chemical components found in licks might be acquired in smaller areas of exposed soil throughout an individual's range, but the large licks in the Tuchodi watershed concentrate multiple chemical components. Hence, the distribution of licks is important in the movements and distribution of ungulate populations.
In summary, this is the 1st study to our knowledge to compare the chemical composition of 2 different types of licks (wet and dry), each used by 2 different ungulate species in the same watershed. We used rigorous sampling techniques and chemical extractions that were adjusted to pH levels of the ungulate digestive tract where elements are absorbed. We approached the system using an animal context to interpret lick use. Our findings show that even though the 2 types of licks are visually distinct, with different concentrations of chemical components, they serve similar functions. Sodium and sulfate concentrations were typically higher at both wet and dry licks than at control sites. Carbonates and magnesium were elevated in soils from dry licks and magnesium also was high in inflow waters to wet licks. We combined our chemical assessments of lick soils with chemical assessments of diets and fecal samples. Forage plants used by all 4 ungulate species were too low in sodium to meet requirements. The high levels of potassium in spring or early-summer forage could compromise absorption of marginal magnesium concentrations in the diet. Fecal samples indicated that animals were in low sodium balance, but because of potentially compromised absorption, they were probably not reflective of an animal's magnesium balance. The licks in the Tuchodi watershed, both wet and dry, appear to provide ungulates with supplemental sources of sodium, carbonates, and magnesium, which cannot be met by foraging strategies alone. In our study, we also retained the clay fraction of the lick samples. The presence of expandable clays was not detected in the few samples analyzed, but further analyses of clay in lick soils should be pursued. The clay content of soils from both wet and dry licks ranged from approximately 50% to 60%. Further studies are needed to explore the role of clay in the foraging strategies of herbivores relative to improving palatability and digestibility (Diamond 1999; Hui 2004; Krishnamanj and Mahaney 2000), especially considering that the use of licks by moose and mountain goats later in the summer (Ayotte 2004) coincides with increased consumption of forages high in plant defense compounds.
Acknowledgments
Funding for this research was provided by the Muskwa-Kechika Trust Fund and the University of Northern British Columbia's Northern Land Use Institute. We acknowledge logistical support from D. Gunn, R. Peck, and L. and L. Warren, and the field assistance of A. Anderson, P. Hirshfield, and M. Shook. Advice on soil analytical methods was provided by D. Dick, P. Sanborn, M. Rutherford, C. Dawson, C. Chisolm, W. Hines, A. Pilley, and G. Odongo. We also are grateful for the ecological contributions from D. Heard.
Literature Cited
Author notes
Associate Editor was Floyd W. Weckerly.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}